Ia from primaries have a powerful excitatory effect on motoneurons of the same muscle and its synergists in adjacent spinal segments (connections may be monosynaptic or polysynaptic). Ia afferents also inhibit alpha motoneurons of antagonistic muscles via inhibitory interneurons and corresponding contralateral muscles. Ia afferents also have a weak polysynaptic excitatory action on gamma motoneurons.
Group II afferents from spindle secondaries also excite alpha motoneurons via mono & polysynaptic paths.
Classical stretch reflex: 'the capacity of a muscle to resist extension' is the result of all these spindle projections to motoneurons and thereby to muscle. The monosynaptic Ia component is responsible for the 'tendon jerk'. The 'tonic stretch reflex' is mainly disynaptic or polysynaptic.
As spinal interneurones are an important target for supraspinal control and gating of reflexes, the tonic stretch reflex is modulatable.
Click on the The monosynaptic reflex in man tab above to continue.
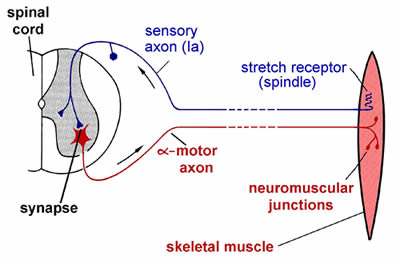
This diagram shows the main components in the simplest kind of reflex, as exemplified by the knee jerk.
If you tap the patellar ligament at the front of the knee with a small tendon hammer, this will cause a short sharp stretch of the muscle that inserts into this tendon.
When the muscle is stretched, the Ia afferents from the spindle are depolarised by the stretch. This generator potential triggers APs which then travel rapidly along the axon of the neuron into its central terminals in the spinal cord.
Some of these central terminals make synaptic contact directly with the cells bodies of motor neurons innervating the muscle that was stretched.
These synapses are excitatory, initiating APs which are propagated via the peripheral axon to its terminals on the muscle fibres, which are in turn are excited, and this stimulates the muscle to contract, counteracting the initial stretch.
As there is only one synapse involved in the CNS pathway here, this is called a mono-synaptic reflex. It has all the basic constituents of a reflex: sensor (in this case themechanosensitive Ia afferent terminals wrapped around the intrafusal fibres), afferent pathway, integrating centre (in this case just the motoneuron cell body), efferent pathway, and effector (muscle in this case). Because both afferent and efferent pathways consist of fast conducting axons (large diameter and myelinated), and there is only one central synapse, this reflex is extremely fast. A very similar monosynaptic reflex response to stretch exists for all muscles that have muscle spindles (there are a very few that don’t), and often the generic term for this kind of monosynaptic muscle reflex is ‘tendon jerk’.
Many reflexes are more complicated than this, involving interneurons in the pathway between afferent and efferent. Reflexes may also be inhibitory; for example, central axons of the same spindle afferents that directly excite the motoneurons innervating this muscle (i.e. the ‘homonymous’ motoneurons in neurophysiologist terms), will also make synaptic contact with interneurons that, when excited, have an inhibitory effect on motoneurons innervating antagonist muscles.
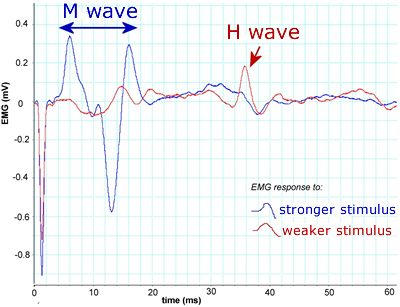
H reflex
To see this, stimulate the nerve to a leg muscle and record EMG. On stimulating a muscle nerve at low stimulus strength (insufficient to activate motor axons directly) a long latency (delayed) muscle response is obtained. This Hoffmann or H reflex is due to excitation of some Ia afferents in the nerve, which then elicit activation of the motoneurons via the monsynaptic reflex. A stronger stimulus strength adds a short latency (early) muscle response due to direct activation of axons of alpha motoneurons at the stimulus point (M response). Further increases in stimulus strength increase the M response and decrease the H response. The decrease in H response occurs because action potentials propogate antidromically towards motoneuron cell bodies and collide with action potentials activated in the alpha motoneurons via the monosynaptic reflex pathway.
These traces show EMG recordings from the lateral gastrocnemius muscle. The muscle nerve (tibial nerve) was stimulated at the back of the knee with 0.5 ms duration square wave current pulse stimuli of gradually increasing size. The red trace shows the response to a relatively weak stimulus. The main response here is a long latency (at ca 35-40 ms) response called the H wave. As stimulus strength increased, responses such as the blue trace were recorded. There is a much larger and shorter latency (at ca 5-15 ms) M wave, and the H wave has almost disappeared.
Jendrassik manoeuvre
The reflex, e.g. a tendon jerk is reinforced (a bigger muscle response) by clenching fists or jaw. This happens because of a general increase in motoneuronal excitability, which means that the same afferent input elicited by stretching the tendon is able to bring a greater proportion of the motoneurons to threshold, enabling them to fire action potentials.
Central Control of Dynamic & Static Fusimotor Neurones
Dynamic and Static fusimotor neurones are under separate central control. Fusimotor activity can be modified by stimulation of many areas of the CNS.
There are separate corticospinal dynamic and static fusimotor pathways. Static gamma motorneurons can be excited or inhibited from many sites in the central nervous system e.g. lateral vestibular nucleaus. Dynamic gamma motorneurons can be excited from an area posterior to the Red Nucleus (the so-called MC area for dynamic control).
Click on the Alpha-Gamma linkage tab above to continue.
Evidence has been obtained to establish that co-activation of alpha and gamma motorneurons happens.
We do not know anything about the time course and intensity of fusimotor outflow in the full range of human movements but can state that during a voluntary contraction alpha and gamma motorneurons are active at the same time. On the otherhand, in freely moving cats gamma motorneurons are active independently of alpha motorneurons under some circumstances.
See 'Stretch Reflexes: Example in Cats' [Opens in new window]
Click on the Planned movements tab above to continue.
It is thought that to make a planned movement, two messages are required:
- A message to the spindle signalling the desired trajectory of movement.
This takes the form of a specific fusimotor drive, appropriate to ‘set’ the spindle afferent firing during the desired movement
- A signal to extrafusal muscle that takes into account any load and fatigue. This takes the form of a specific frequency of stimulation delivered to an appropriate proportion of motoneurons supplying the muscles in question – designed to produce a particular force.
If the load proves unexpectedly great, the spindle will suffer a smaller degree of unloading because the muscle is shortening less than expected. As a result, afferent discharge will increase (exciting additional motoneurons) and movement will be reflexly assisted. The discharge may also cause a central command to increase motor outflow.
Click on the Conscious appreciation of position tab above to continue.
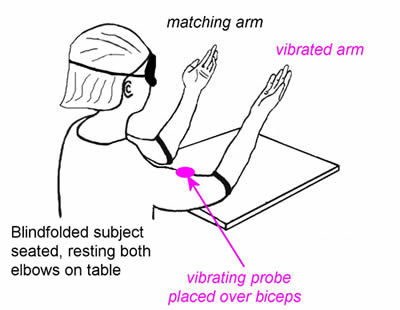
Spindles contribute to proprioception (sense of position) and to kinaesthesia (sense of movement).
This can be easily demonstrated by vibration of muscle tendons, which selectively excites primary endings and causes large position errors in a blindfolded subject.
The secondary sensory endings provide a much better length signal than the primary ending, but in this experiment the vibration so powerfully excites Ia endings that although the sense of position is not lost, the length signal is wrongly calibrated.
The experimenter moves the blindfolded subject’s ‘test’ arm to a series of positions, and asks him/her to match those positions with the other arm. Most people can match the position very well, with little error. However, if the experimentor repeats the experiment while there is a vibrating probe over biceps on the test arm, subjects make quite substantial errors when trying the match its position with the other arm.
Select another item from the menu on left to continue.